
Intens træning aktiverer proteinsyntese, men kun hvis den rigtige ernæring er der for at understøtte den. Hvis du læser artiklen John Meadows og jeg skrev kaldet Maximize Protein Synthesis, eller hvis du har læst noget, som T Nation har sagt om emnet i årevis, er du sandsynligvis bekendt med begrebet ”det anabolske vindue” og vigtigheden af peri-træning ernæring.
Hvad der sker på mobilniveau i timerne efter træning, forudsiger langsigtede gevinster. Udnyt dette “anabolske vindue”, så vokser du som aldrig før. Gå glip af det konsekvent, og held og lykke.
At få de rigtige makronæringsstoffer til de rigtige tidspunkter er nøglen, men makroer er kun en del af det store billede. Et vigtigt, men ofte overset aspekt af muskelproteinsyntese er cellevolumen. Cellevolumen er ikke kun kosmetisk; det er den vigtigste drivkraft for aminosyretransport og arbejder også bag kulisserne for at aktivere proteinsyntese og undertrykke proteinnedbrydning.
En fuld / volumineret muskel er en anabolsk muskel. Mens vi har kendt i over 20 år, at celle hævelse hæmmer proteinsnedsættelse og stimulerer proteinsyntese i visse celler (1-3), var mekanismen, der forbinder cellevolumen til proteinsyntese, indtil for nylig et mysterium.
Hvad vi ved nu er, at proteinsyntese styres af enzymet mTOR, som aktiveres af mekanisk stress, vækstfaktorer og leucin.
Mens alle disse tre er vigtige for træningsstimuleringen, er mTOR-signalering også afhængig af cellevolumen.(4) Dette er især vigtigt i skeletmuskulatur, hvor cellevolumisering aktiverer glykogen- og proteinsyntese og hæmmer proteinnedbrydning.(5, 6)
Det videnskabelige gennembrud, der førte til sammenhængen mellem cellevolumen og proteinsyntese, fandt sted i 2005, hvor en gruppe forskere opdagede, at det kræves mere end bare leucin for at aktivere mTOR - glutamin er også påkrævet.(7)
Dette kom som en overraskelse. Selvom glutamin betragtes som en "betinget essentiel" aminosyre, der begrænser nedbrydning af protein under alvorligt traume eller stress, havde det aldrig været forbundet med mTOR-aktivering.
Glutamin var nødvendigt for leucinoptagelse og cellevolumisering, som begge er nødvendige for at aktivere proteinsyntese. Forfatterne fortsatte med at vise den cellulære glutamin udtømning resulterer ikke kun i en reduktion af cellevolumen, men reducerer også leucins evne til at aktivere proteinsyntese.(7)
Denne opdagelse var enorm, fordi den gav en direkte forbindelse mellem glutamin, cellevolumisering og proteinsyntese. For første gang blev det vist, at glutamin er nødvendig for aktivering af proteinsyntese med leucin.
Mens denne undersøgelse antydede, at glutamin er et meget vigtigt stykke af puslespillet, der forbinder cellevolumen og proteinsyntese, blev den nøjagtige mekanisme ikke udarbejdet før 2009, da Nicklin et al. opdagede, at eksport af glutamin er koblet til leucinimport og mTOR-aktivering.(8)
For at få leucin ind i cellen er der en indledende periode med ”glutaminbelastning.”Dette trækker også vand ind, hvilket øger cellevolumenet. Efter "glutaminbelastningsfasen" eksporteres glutamin ud af cellen til gengæld for import af leucin.
Nicklin et al. opdagede også, at cellulære glutaminniveauer er hastighedsbegrænsende for aktivering af proteinsyntese med leucin. Når celler blev behandlet samtidigt med glutamin og en EAA-blanding indeholdende leucin, blev aktivering af proteinsyntese forsinket i 60 minutter. Når de samme celler var glutamin "forudindlæst", blev proteinsyntese tændt inden for 1-2 minutter efter, at cellerne fik leucin.
Dette resultat var vigtigt, fordi det forklarede forsinkelsestiden for aktivering af proteinsyntese med leucin i denne eksperimentelle model.
Disse resultater kaster endelig lidt lys over det cellulære maskineri, der regulerer aminosyretransport, og hvordan det er koblet til kontrol af proteinsyntese.
Dette arbejde skal dog fortolkes med en vis forsigtighed. Et vigtigt forbehold for disse undersøgelser var, at de blev udført in vitro (i.e. i cellekulturer) hvor regulering eller proteinsyntese er meget enklere. Muskelceller er i stand til at fremstille glutamin efter behov fra andre aminosyrer, og "glutaminudtømning" i denne cellekulturmodel er ikke repræsentativ for mere fysiologiske situationer in vivo.
Høje proteinsyntesehastigheder kan ikke opretholdes på ubestemt tid i muskelvæv med eller uden glutamintilskud. Glutamin kan dog bruges til strategisk at understøtte proteinsyntese ved at optimere cellevolumisering i løbet af perioden efter træning.

Celler har meget travlt, og der er mange membranbundne ionkanaler og transportørproteiner, der regulerer trafik ind og ud af cellen. Der er især to klasser af aminosyretransportører, der er vigtige her: "System L" og "System A" aminosyretransportører er tætest knyttet til mTOR-signalering og proteinsyntese.(8-10)
Aktiviteten af System A- og System L-transportører er koblet, hvilket gør det muligt for leucin og de andre BCAA'er at blive absorberet i cellen.(11) System L-transportører er ansvarlige for tilstrømningen af leucin og de andre BCAA'er til gengæld for udstrømningen af glutamin.
System A-transportører arbejder dog gennem en anden mekanisme, hvor glutamin er koblet til natriumoptagelse.(12, 13) Koblingen mellem natriumoptagelse og System L / System A-aminosyretransportører kaldes Tertiær Aktiv Transport (forkortet TAT). Det er TAT, der i sidste ende driver leucin inde i cellen, hvilket fører til mTOR-aktivering og proteinsyntese.(11)
Du kan se, hvordan TAT fungerer i nedenstående figur:
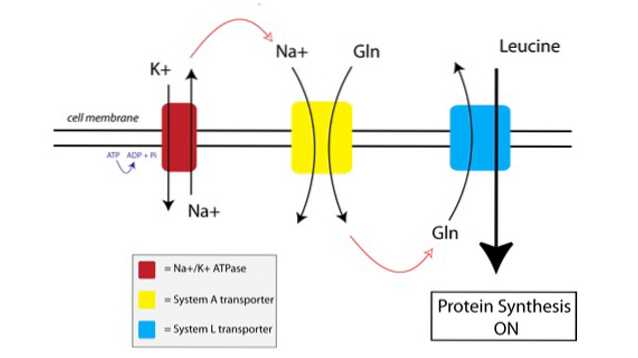
For det første bruger en membranbundet pumpe kaldet natrium-kalium ATPase-pumpen (Na + / K + ATPase, rød i figuren ovenfor) energi fra ATP til at flytte natrium uden for cellen mod dets koncentrationsgradient.
Den øgede koncentration af natrium uden for cellen er koblet til importen af glutamin af System A-transportøren (gul i figuren). Glutamin og natriumtilstrømning til cellen trækker også ekstra vand ind, hvilket får cellen til at svulme op. Dette sætter cellen i en anabol tilstand, hvorved proteinsyntese-maskinerne startes til aktivering.
Når glutamin opbygges til tilstrækkeligt høje niveauer inde i cellen, aktiveres System L-transportører (blå i figuren), som skifter glutamin uden for cellen i bytte for leucinoptagelse. Indgang af leucin i cellen er udløseren til proteinsyntese.
Mens dette hidtil har været en stor lektion i biokemi, er opdagelsen af TAT ikke kun vigtig for cellebiologer. Nu hvor vi ved, hvordan cellevolumen er koblet til aminosyretransport og proteinsyntese, kan vi designe flere ernæringsstrategier for at maksimere processen, når den tæller - i den kritiske periode efter træning.
Proteinsyntese er helt afhængig af cellehydrering - hvis du endda er lidt dehydreret, er evnen til at komme sig efter intens træning totalt kompromitteret. At få masser af vand er ikke en god idé her, men vand alene er ikke nok.
Elektrolytter som natrium, kalium, chlorid og fosfat fungerer også som ”osmolytter”, fordi de trækker vand ind i cellen. Efter en intens træningssession har vi brug for vand, aminosyrer og elektrolytter for at maksimere cellevolumiseringsprocessen, der driver proteinsyntese.
Natrium, magnesium, calcium, kalium, fosfat og chlorid (for at nævne nogle få) er alle vigtige her. På et minimalt niveau (medmindre du gør det under råd fra din læge), skal du ikke vige tilbage for natrium før eller efter træning. Hvis du er natriumforarmet, vil din pumpe fra træning næsten ikke være eksisterende, og natrium er nødvendigt for glutaminoptagelse.
For at eliminere gætteriet er Surge® Workout Fuel og Plazma ™ designet med ideelle elektrolytforhold til understøttelse af cellevolumen og proteinsyntese.
Glutaminoptagelse i cellen forårsager cellevolumisering, hvorved muskelceller primeres til proteinsyntese. Som nævnt er en fuld / volumineret muskel en anabolsk muskel. Sammen med kørsel af aminosyretransport øger cellevolumisering også glykogensyntese og hæmmer proteinnedbrydning.(4-6)
Proteinsyntese undertrykkes af nedbrydning af glutamin, som har enorme konsekvenser for atleter med hård træning. Efter en intens træning er der monteret en inflammatorisk reaktion, som gør det muligt for immunceller at komme ind i stødt muskelvæv for at starte reparations- / genopbygningsprocessen.(14)
Glutamin optages så hurtigt af immunceller, at det betragtes som ”brændstof i immunsystemet.”(15) Ikke overraskende er det blevet vist, at intens træning forårsager plasmaglutaminudtømning.(16-18)
Af denne grund øges glutaminbehovet i perioden efter træning, hvor det lokale immunrespons kan konkurrere om tilgængeligheden af glutamin til primære muskelceller til aminosyreoptagelse og proteinsyntese.
Forudindlæsning celler med glutamin kan også potentielt reducere "forsinkelsestid" forbundet med leucinaktivering af proteinsyntese. Hvis du ikke allerede gør dette, skal du tage 10-15 g glutamin eller glutaminpeptider straks efter træning. Fordi BCAA'er er et andet yndlingssubstrat til muskelglutaminsyntese og det også har vist sig at øge muskelglutaminproduktionen (19-21), er BCAA'er og leucin også nyttige i perioden før træning for at hjælpe med at maksimere endogen produktion af glutamin.
Redaktørens bemærkning: Hele Plamza / MAG-10-protokollen med dens træning, træning og ernæring efter træning opfylder forfatterens anbefalinger til belastning af musklerne med BCAA'er og leucin.
For nylig blev det opdaget, at forbruget af EAA'er øger ekspressionen af både System A og System L-aminosyretransportører.(9) Det er vigtigt, at dette sker på "post-transkriptionelt niveau", hvilket betyder på niveauet med proteinsyntese, hvor eksisterende mRNA'er oversættes til proteiner.
Kontrast dette med “denovo”Proteinekspression - hvor det kan tage 16 eller flere timer at syntetisere, behandle og transportere nye mRNA'er - post-transkriptionel aktivering af proteinsyntese kan forekomme i minutter til timer, så celler hurtigt kan øge niveauet af bestemte proteiner, når det er nødvendigt.
Pludselig har vi mere incitament til at have en solid ernæringsplan for peri-træning - EAA-indtag i præ- og peri-træningsperioderne betaler sig stor post-træning ved at øge ekspressionen af aminosyretransportører, hvorved cellen primeres for maksimal aminosyreoptagelse og aktivering af proteinsyntese.
Hurtigt absorberende proteinisolater eller hydrolysater såsom Mag-10® Protein Pulsing Protocol ™ eller Plazma ™ i perioderne før og peri-træning er ideelle her.
Insulin er det mest anabolske hormon i kroppen. Sammen med direkte aktivering af proteinsyntese øger insulin også translokation af System A-aminosyretransportører til cellemembranen.(22)
Dette betyder, at insulin får flere System A-transportører til at blive vist på cellemembranen, klar til at drive mere glutamin ind i cellen. Mere glutamin fører til mere cellevolumen, hvilket driver mere leucin ind i cellen, hvilket i sidste ende fører til mere proteinsyntese.
Mens EAA'er øger udtryk af AA-transportører, er det insulinsignalet, der gør det muligt for dem at blive vist på celleoverfladen, klar til at skifte nye aminosyrer ind i cellen.
Dette er endnu en grund til, at præ-og peri-træningskulhydrater er en god idé, medmindre du er i ekstrem fedtreduktionstilstand: insulin øger kapaciteten til cellulær aminosyretransport.
Kulhydrater øger insulinniveauet, men visse aminosyrer kan også bruges til at forstærke frigivelsen af insulin. Glutamin er en kraftig aktivator af "incretin" hormoner, som gør insulinproducerende celler i bugspytkirtlen mere følsomme over for glukose.(23) Glycin forstærker også frigivelse af insulin gennem en anden mekanisme.
Selvom kulhydrater efter træning alene øger insulinniveauerne, vil kombination af disse insulin-potentierende aminosyrer med kulhydrater overbelaste din bugspytkirtlen for endnu mere frigivelse af insulin. Selvom det er godt at holde insulinniveauerne på den nedre side det meste af tiden, maksimerer øgede insulinniveauer i peri-træningsperioden aminosyretransport, cellevolumen og proteinsyntese og undertrykker også nedbrydning af protein.
Den type intens anaerob træning, der kræves for at opbygge masser af kvalitetsmuskel, fører til betydelig lactatproduktion, faldende muskel-pH. Dette fører til tidlig muskeltræthed og svaghed, men også visse aminosyretransportører, inklusive system A, hæmmes af lav pH.(13)
Når muskel-pH er lav, reduceres optagelsen af aminosyrer, hvilket undertrykker mTOR-aktivering af proteinsyntese.(24) Det er også blevet påvist, at lav pH-hæmning af System A-aminosyretransportører øger proteinnedbrydning.(25)
Det er her beta-alanin kommer ind. Øgede muskelcarnosinniveauer fungerer som en naturlig syrebuffer, der udvider den anaerobe tærskel ved at begrænse faldet i muskel-pH fra træning.
Beta-alanin har en anden vigtig funktion: at hjælpe med at opretholde proteinsyntese og få det online hurtigere efter intens træning ved at forhindre dæmpning af aminosyretransport.
For at øge muskelcarnosinniveauet skal du tage to tabletter Beta-7 ™ tre gange om dagen.
Under intense træningssessioner reduceres proteinsyntese, og proteinnedbrydning aktiveres. Dette er uundgåeligt for enhver hård træningsløfter. Det omfang, i hvilket vi kan minimere de katabolske virkninger af træning, og jo hurtigere vi kan vende tilbage til "anabolsk tilstand" i perioden efter træning bestemmer i sidste ende, hvor effektivt vi vil komme - og vokse.
Makronæringsstoffer er vigtig, men det er et middel til et mål. Cellevolumen er den vigtigste drivkraft for aminosyretransport og proteinsyntese. Ved at forstå, hvordan aminosyretransport sker, og hvordan det reguleres af cellevolumen, kan vi hurtigere få mere leucin ind i trashede muskelceller og derved ryge den anabolske ild og i sidste ende føre til bedre gevinster.
Ovenstående strategier er effektive, praktiske og er baseret på den nyeste videnskabelige forskning. Brug dem som en skabelon til at tage din træningsfremgang til det næste niveau.
Jeg ser frem til dine spørgsmål om LiveSpill!



Endnu ingen kommentarer